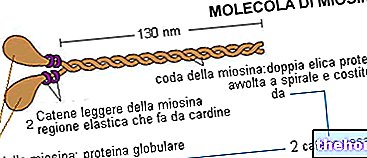
Tack vare dessa cylindriska enheter omvandlas den kemiska energin som frigörs genom metaboliska reaktioner till mekanisk energi; genom att sätta in sig själv genom senorna och verka på benhävarmarna genererar muskeln rörelse.
Skelettmuskelfibrer varierar i längd från några millimeter till flera centimeter, med en diameter från 10 till 100 µm (1 µm = 0,001 mm); de är de största cellerna i kroppen.
"Cytologiskt sett" är fiberceller resultatet av en process som kallas myogenes, som är sammansmältningen av flera myoblaster - en åtgärd som är beroende av muskelspecifika proteiner som kallas fusogener, myomaker eller myomerger. Det är därför som myocellerna framstår som långa cylindriska och polynuklerade celler (som innehåller många myonuclei - bland annat tydligt synliga på ytan under mikroskopet).
En muskelfiber, t.ex. i brachial biceps, med en längd av 10 cm kan den ha upp till 3000 kärnor.
Inuti dem finns det istället tusentals filament, kallade myofibriller, som innehåller kontraktila enheter som kallas sarkomerer.
Fysiologer som hanterar muskler berättar att de olika fibrerna skiljer sig från varandra, inte bara ur anatomisk synvinkel, utan också för några exakta fysiologiska egenskaper.
Därför känner man igen olika typer av fibrer inom varje muskel, klassificerade enligt olika kriterier som energimetabolism, kontraktionshastighet, motståndskraft mot trötthet, färg, etc.
Sammantaget är en enda muskel som t.ex. brachial biceps, cirka 253 000 muskelfibrer finns.
Visste du att ...
Mellan basalmembranet och muskelfibrernas sarkolemma ligger en grupp muskelstamceller som kallas myosatellitceller.
Dessa är normalt vilande men kan aktiveras av träning eller sjukdom för att ge ytterligare myonuclei som behövs för muskeltillväxt eller reparation.
specifika, fosfager (ATP och CP), mitokondrier, myoglobin, glykogen och en högre kapillär densitet.
Muskelceller kan dock inte dela sig för att producera nya celler och som ett resultat deras antal tenderar att minska med åldern.
), som ger upphov till tre typer av fibrer.
Dessa fibrer har relativt distinkta metaboliska, kontraktila och motoriska egenskaper - sammanfattade i tabellen nedan.
VIKTIG! De olika egenskaperna, även om de delvis beror på de enskilda fibrernas egenskaper, tenderar att vara mer relevanta när de mäts på motorenhetens nivå - vilket dock visar mycket minimala variationer vad gäller olika fibrer - snarare än av den enda fibern.
Låt oss nu se några typer av klassificering.
Fiberfärg
Traditionellt klassificerades fibrer efter deras färg, vilket beror på myoglobininnehållet.
Typ I -fibrer verkar röda på grund av höga myoglobinnivåer, tenderar att ha mer mitokondrier och högre lokal kapillartäthet.
De är långsammare att krympa men mer lämpade för resistens, eftersom de använder oxidativ metabolism för att generera ATP (adenosintrifosfat) från glukos och fettsyror.
De mindre oxidativa typ II -fibrerna är vita eller i alla fall klara på grund av bristen på myoglobin och koncentrationen av glykolytiska enzymer.
Kontraktionshastighet
Fibrer kan klassificeras enligt deras kontraktila hastigheter i snabba och långsamma. Dessa egenskaper överlappar till stor del, men inte helt, med klassificeringar baserade på färg, ATPase och MHC.
- Fibrer a snabb sammandragning de där myosin kan dela ATP mycket snabbt. Dessa inkluderar typ II ATPase och typ II MHC -fibrer. De visar också en större kapacitet för elektrokemisk överföring av åtgärdspotentialer och en snabb nivå av kalciumfrisättning och absorption av det sarkoplasmatiska retikulumet.De är baserade på ett väl utvecklat, anaerobt, snabbt energiöverföringsglykolytiskt system och kan dra ihop sig 2-3 gånger snabbare än långsamma ryckfibrer Snabba ryckmuskler är lämpliga för att generera korta utbrott av styrka eller hastighet än långsamma muskler, och därför trötthet snabbare.
- Fibrer a långsam kontraktion genererar energi för resyntesen av ATP genom ett aerobt och långvarigt överföringssystem. Dessa inkluderar främst fibrer av typ ATPase typ I och MHC typ I. De tenderar att ha en låg nivå av ATPas -aktivitet, en långsammare ryckhastighet med en mindre utvecklad glykolytisk kapacitet. Långsamma ryckfibrer utvecklar mer mitokondrier och kapillärer, vilket gör dem bättre för uthållighetsarbete .
Fiber typmetoder
Det finns ett antal metoder som används för fibertypning, vilket ofta skapar viss förvirring bland icke-experter.
Två ofta otvetydiga metoder är histokemisk färgning för myosin ATPas -aktivitet och immunhistokemisk färgning för myosin -tungkedjetyp (MHC).
Aktiviteten för myosin ATPase -enzymet kallas vanligen och korrekt helt enkelt som "fibertyp" och härrör från den direkta mätningen av aktiviteten hos ATPase -enzymet under olika förhållanden (t.ex. pH).
Myosin -tungkedjefärgningen kallas mer exakt som "MHC -typ" (myosin tung kedja) och, som kan förstås, resultat från bestämning av olika MHC -isoformer.
Dessa metoder är fysiologiskt besläktade, eftersom MHC -typen är den huvudsakliga determinanten för ATPas -aktivitet. Ingen av dessa skrivmetoder är emellertid direkt metabolisk. det är de tar inte direkt upp fiberns oxidativa eller glykolytiska kapacitet.
När man hänvisar till "typ I" eller "typ II" fibrer, refererar detta mer exakt till bedömningen genom färgning av "ATPas -aktiviteten för myosin (t.ex." typ II "-fibrer hänvisar till typ IIA + typ IIAX + typ IIXA ... etc.).
Nedan visas en tabell som visar sambandet mellan dessa två metoder, begränsat till de typer av fibrer som finns hos människor.Subtypens versaler används vid fibertypning jämfört med MHC -typning; vissa typer av ATPase innehåller faktiskt flera typer av MHC.
Vidare uttrycks inte en undertyp B eller b hos människor med någon av metoderna. Tidiga forskare trodde att människor kunde uttrycka en MHC IIb, vilket ledde till ATPas -klassificeringen av IIB. Efterföljande forskning har dock visat att humant MHC IIb faktiskt är IIx, vilket indikerar att den mer korrekta formuleringen är IIx.
Subtyp IIb eller IIB, IIc och IId uttrycks istället i andra däggdjur, vilket är allmänt dokumenterat i litteraturen.
Ytterligare fiberskrivningsmetoder beskrivs på ett mindre formellt sätt och finns på fler spektra, till exempel den som normalt används inom friidrott.
De tenderar att fokusera mer på metabolisk och funktionell kapacitet (kontraktionstid, främst oxidativ vs anaerob mjölksyra vs. anaerob mjölksyra, snabb kontra långsam kontraktionstid).
Som nämnts ovan mäter eller dikterar inte fibertypning med ATPase eller MHC dessa parametrar. Många av de olika metoderna är dock mekaniskt kopplade, medan andra är relaterade in vivo.
T.ex, typen av ATPase -fiber är relaterad till sammandragningshastigheten, eftersom den höga aktiviteten hos ATPase möjliggör en snabbare cykel av tvärbryggan. Typ I -fibrer är "långsamma", delvis eftersom de har låga ATPas -aktivitetshastigheter jämfört med typ II -fibrer, men att mäta kontraktionshastigheten är inte detsamma som att skriva ATPas -fibern.
, vita och mellanliggande fibrer. Deras proportioner varierar dock beroende på det fysiologiskt tilldelade arbetet till den muskeln.Till exempel, hos människor, innehåller quadriceps -musklerna cirka 52% av typ I -fibrer, medan soleus är cirka 80%. Orbicularis -muskeln i ögat har däremot bara cirka 15% av typ I.
Visste du att ...
Kraften som utvecklas av en muskelfiber beror på dess längd i början av kontraktionen. Det måste ha ett optimalt värde, utanför vilket (indragna eller alltför sträckta muskler) styrka prestanda reduceras. När det gäller muskelförstärkning är det vanligaste misstaget att arbeta musklerna redan i delvis förkortning. De enda undantagen från regeln är närvaron av smärta eller obehag, eller paramorfismer, som därför kräver en begränsning av rörelseområdet (ROM).
De övervägande vita musklerna, rika på typ II -fibrer, kallas fasiska, eftersom de kan snabba och korta sammandragningar. De röda musklerna däremot, där typ I -fibrer råder, kallas tonic, på grund av förmågan att stanna kvar i sammandragning under lång tid.
De motoriska enheterna i muskeln visar dock mycket liten variation, vilket gör att dimensionell princip för rekrytering av motorenheter; det vill säga, beroende på intensitet / styrka som krävs, kan kroppen bara stimulera vissa (t.ex. vid långvarig aerob aktivitet) eller alla (t.ex. under en maximal knäböj) enheterna i fråga.
Idag vet vi att det inte finns några könsrelaterade skillnader i fördelningen av fibrer. Emellertid kan proportionerna av de olika typerna - som vi vet varierar mycket mellan djurarter och i mindre utsträckning mellan etniciteter - "kunna" variera avsevärt från person till person.
Enligt vissa insikter bör stillasittande män och kvinnor (liksom små barn) ha 55% fiber av typ I och 45% typ II -fiber.
Höga idrottare, å andra sidan, har en specifik fiberfördelning baserad på vilken typ av metabolism som används. Längdåkare har huvudsakligen fibrer I, sprinters främst II och medeldistanslöpare, kastare och hoppare, nästan överlappande procentandelar av båda.
Det har därför föreslagits att olika typer av träning kan framkalla signifikanta förändringar i skelettmuskelfibrer, även om det inte är möjligt att med säkerhet fastställa vad den redan existerande genetiska sammansättningen för samma ämnen var. Denna process "kan" tillåtas av fibrernas specialiseringskapacitet, eller till och med bara en del, som tillhör makrouppsättning II.
Det är möjligt att fibrer av typ IIx visar förbättringar i oxidativ kapacitet efter högintensiv uthållighetsträning, vilket leder dem till en nivå där de skulle kunna uppfylla oxidativ metabolism lika effektivt som fibrer I hos otränade ämnen.
Detta skulle bestämmas av en ökning av storleken och antalet mitokondrier och deras associerade förändringar, men inte av en förändring i typ av fiber..