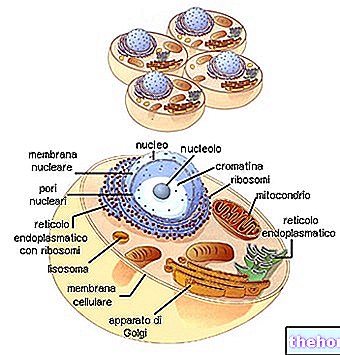
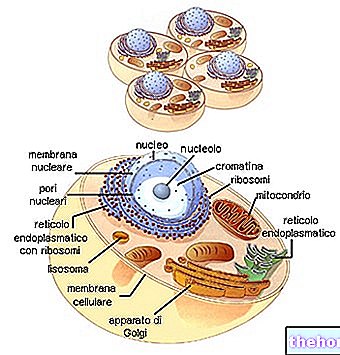
Levande varelsers cykliska kontinuitet hittar länkarna mellan på varandra följande generationer i reproduktionens fenomen.
Reproduktion utförs på olika nivåer av den evolutionära skalan, i de olika förgreningarna av grönsaks- och djurriken, i de olika levande arterna, med en sådan mängd olika mekanismer som ensam motiverar en hel avhandling.
En första klassificering av reproduktionsfenomen måste skilja encelliga från flercelliga organismer, eftersom endast i den förra sammanfaller celldelning med reproduktion.
I flercelliga kan reproduktion vara agamisk eller sexuell (eller gamic).
Agamisk reproduktion, relativt mindre frekvent, baseras på mitosmekanismen, så att artens variabilitet snarare anförtros upprepade förekomster av mutationer.
Det finns också olika mekanismer, såsom strobilisering, sporulation, etc., medan vi i växter hittar regenerativa former välkända inom jordbruket (sticklingar, skiktning etc.).
Den mest utbredda reproduktionsmekanismen i de högre formerna är dock den sexuella, motsvarande utseendet på meios, bildandet av könsceller och deras fusion i zygoten (befruktning).
Hos primitiva arter är könscellerna inte morfologiskt differentierade: i detta fall talar vi om isogametia. Det finns emellertid alltid två serier av könsceller, identifierade med symbolerna (+) och (-) och befruktning kan endast ske från "mötet mellan könsceller med motsatt tecken: det finns därför en biologisk skillnad som ännu inte uttrycks morfologiskt.
I takt med att den evolutionära skalan fortskrider framträder morfologisk och funktionell differentiering, där en typ av kvinnliga könsceller i allmänhet är utrustad med rikligt med reservmaterial (deutoplasma eller kalv, vilket kommer att säkerställa utvecklingen av embryot tills det kan vara metaboliskt oberoende) och en typ av manliga könsceller utrustade med rörlighet för att nå kvinnliga. Gameter är alltid haploida och är resultatet av meios, deras fusion ger upphov till diploida zygoten.
Mellan meíosis och zygote kan en serie haploida cellgenerationer förgå, precis som mellan zygote och meios kan en serie diploida cellgenerationer förgå, med de olika sorterna av generativa cykler uttryckta genom generationsväxling.
En diplonte -art (med en diploid organism) kännetecknas av gametisk meios: meios producerar direkt könscellerna, som genom sammanslagning omedelbart rekonstituerar det diploida tillståndet. Detta är det vanliga fallet med Metazoans, inklusive människan.
Gametogenes
Med tanke på reproduktionen med gametisk meios, som den förekommer hos människan, låt oss försöka klargöra hur meios passar in i gametogenes (bildning av gameter).
Vid embryonisk utveckling, av manlig och kvinnlig gametogenes (kallad spermatogenes och oogenes), finns det en tidig differentiering mellan cellerna som är avsedda att bilda kroppen (somatisk linje) och de som är avsedda att producera könsceller (könslinje). De ursprungliga cellerna i könslinjen kallas protogoner. Med differentiering av gonaden i manlig eller kvinnlig mening finns det differentiering av könscellerna respektive i spermatogoni och ovogonia.
När vi tittar på spermatogenes ser vi att det finns en serie cellgenerationer i spermatogonin, som fortsätter hela livet. Endast en del av spermatogonin som sålunda kontinuerligt produceras skiljer sig från den normala mitotiska cykeln och den meiotiska börjar istället.
Den könscell i vilken meios kommer att börja (reduplikation och sedan första division) kallas första ordningens spermatocyt; dess uppdelning ger upphov till två andra ordningens spermatocyter, som med andra divisionen ger upphov till totalt fyra spermatider.
Vi kan gradera reduktionen av kromosomkittet från 4n av första ordningens spermatocyter (efter reduplikation finns det fyra kromatiler för varje par homologer) till 2n av andra ordningens spermatocyter och till n av spermatiderna, som redan sett genom att studera meios, som Således drar vi slutsatsen: Spermatiderna är därför redan haploida, men de är ännu inte mogna könsceller. Från strukturen hos en cell av haploid typ omvandlar funktionell mognad (kallad spermiohistogenes) spermatiderna till spermatozoer, det vill säga mogna manliga könsceller.
I kvinnlig gametogenes (eller oogenes) finns det flera skillnader. Först och främst är antalet könsceller som ska förberedas mycket mindre. Det uppskattas att cirka 5 X 105 ovogoner framställs i könskörtlarna hos en hona av den mänskliga arten; av dessa är bara cirka 400 intresserade av follikelmognad och efterföljande nedstämdhet, i en cykel som vanligtvis bara påverkar en follikel per månad under en bördig period på cirka 35 år.
Det olika antalet könsceller framställda i de två könen motsvarar skillnaden i funktion och beteende som redan nämnts: spermierna är små, rörliga och många i förhållande till behovet av att söka efter ägget och den låga sannolikheten att hitta det; äggen är stora, inerta och få, i förhållande till funktionen att garantera embryot reservmaterialet och det skydd som de får genom intern befruktning (naturligtvis, särskilt vid extern befruktning måste äggstockarna också vara fler).
Behovet av att förse könsceller med reservmaterial motsvarar närvaron i "oogenesen" av en fas av stopp av meios, under vilken kromosomerna delvis despiraliseras. Vi observerar sedan de så kallade "fjäderkromosomerna", där en serie av extroversions identifierar de egenskaper i vilka generna som är ansvariga för syntesen av deutoplasma är deprimerade.
Det mindre antal som krävs för kvinnliga könsceller motsvarar också det faktum att av de fyra haploida celler som produceras av meios, bara en tar emot hela reservmaterialet och blir en könscykel, medan de andra tre (polocyterna eller polära kropparna), som endast innehåller kromosomalt material, kan inte ge upphov till zygoter och embryon och är avsedda att gå tillbaka.
Befruktning
Befruktning, det vill säga mötet mellan den manliga och kvinnliga könscykeln, kan utföras på mycket olika sätt.I djurriket observerar vi övergången från yttre befruktning (könsceller som utsätts för miljöfaror och därför nödvändigtvis mycket många i de två könen) till intern befruktning, till vilken föräldravården ytterligare är kopplad till däggdjurs metaboliska förhållande mellan mor och foster .
Befruktning, när mötet mellan könsceller av motsatt kön har ägt rum, måste ske och garantera två villkor: specificitet och unikhet. Det vill säga det måste säkerställas att spermierna är av samma art som ägget och att när den första kommer in kommer inga andra in.
Specificiteten säkerställs av de biokemiska egenskaperna hos akrosomen och äggets yta. I själva verket talas det om reaktioner mellan «fertilisiner» och «antifertilisiner», med specificitet jämförbar med mötet mellan enzymer och substrat.
Befruktningens unika karaktär garanteras av en modifiering av äggets ytstruktur («kortikal reaktion») som börjar precis vid den första specifika fertilisin / antifertilisinreaktionen; efter denna reaktion ändras äggets membran, så att alla andra spermier som når det inte längre kan initiera den specifika befruktningsreaktionen.
Efter befruktningen förblir spermotozoans svans yttre än ägget, medan det kromosomala materialet tränger in i det. Detta, kallat "manlig pronucleus", förenar äggets "kvinnliga pronucleus" och bildar således zygotens diploida kärna.