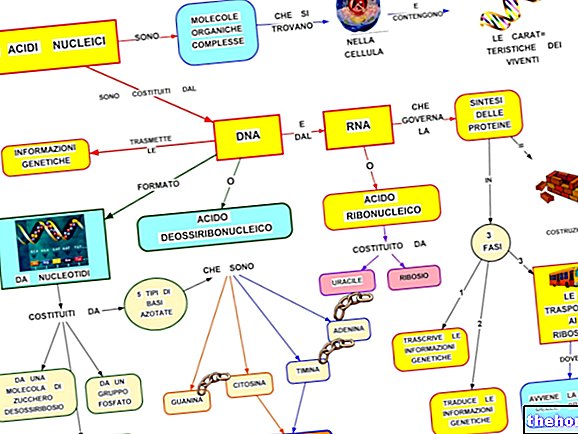
För att det ska finnas en överensstämmelse mellan informationen om polynukleotiden och den för polypeptiden finns det en kod: den genetiska koden.
De allmänna egenskaperna hos den genetiska koden kan listas enligt följande:
Den genetiska koden består av trillingar och saknar inre skiljetecken (Crick & Brenner,).
Det "dekrypterades genom användning av" öppna cellöversättningssystem "(Nirenberg & Matthaei, 1961; Nirenberg & Leder, 1964; Korana, 1964).
Det är mycket degenererat (synonymer).
Organisationen av kodtabellen är inte av misstag.
Trillingar "nonsens".
Den genetiska koden är "standard", men inte "universell".
När man tittar på tabellen med den genetiska koden måste man komma ihåg att den hänvisar till översättningen av "RNAm till polypeptid, för vilka de involverade nukleotidbaserna är A, U, G, C. Biosyntesen för en polypeptidkedja är translationen av nukleotidsekvensen i sekvens aminosyra.

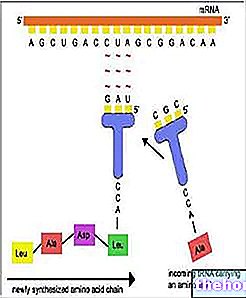
Varje bas -triplett av RNAm, kallad kodon, har den första basen i den vänstra kolumnen, den andra i den översta raden, den tredje i den högra kolumnen. Låt oss ta till exempel tryptofan (dvs. Try) och vi ser att motsvarande kodon vara, i ordning, UGG. Faktum är att den första basen, U, innehåller hela raden med rutor högst upp; i detta identifierar G rutan längst till höger och den fjärde raden i själva rutan, där vi hittar det skrivna Try. På liknande sätt kan vi för att syntetisera tetrapeptiden Leucine-Alanine-Arginine-Serína (symbolerna Leu-Ala-Arg-Ser) hitta kodonerna UUA-AUC-AGA-UCA i koden.

Vid denna punkt bör det dock noteras att alla aminosyror i vår tetrapeptid kodas (till skillnad från tryptofan) av mer än ett kodon. Det är ingen slump att vi i det nyss rapporterade exemplet har valt de angivna kodonerna.Vi kunde ha kodat samma tripeptid med en annan RNAm-sekvens, såsom CUC-GCC-CGG-UCC.
Ursprungligen fick det faktum att en enda aminosyra motsvarade mer än en trilling en betydelse av slumpmässighet, också uttryckt i valet av kodens degenerering, som används för att definiera fenomenet synonymi. Å andra sidan tyder vissa data på att tillgängligheten av synonymer som hänför sig till olika stabilitet i den genetiska informationen inte alls är av misstag.Detta verkar också bekräftas genom att man har funnit ett annat värde för A + T / G + C -förhållandet i de olika utvecklingsstadierna. Till exempel, i prokaryoter, där behovet av variabilitet inte tillgodoses av reglerna för mendelism och neo-mendelism, tenderar A + T / G + C-förhållandet att öka. Den därmed lägre stabiliteten, inför mutationer, ger större möjligheter för variationer slumpmässigt från genmutation.
I eukaryoter, särskilt i multicellulära celler, där det är nödvändigt att cellerna i den enda organismen alla behåller samma ärftliga arv, tenderar A + T / G + C -förhållandet i DNA att minska, vilket minskar möjligheten för somatiska genmutationer .
Förekomsten av synonyma kodoner i den genetiska koden väcker redan nämnda problem med antikodonernas mångfald eller inte i RNAt.
Det är säkert att det finns minst en RNAt för varje aminosyra, men det är inte lika säkert om en enda RNAt kan binda till ett enda kodon, eller kan känna igen synonymer likgiltigt (särskilt när dessa skiljer sig endast för den tredje basen).
Vi kan dra slutsatsen att det i genomsnitt finns tre synonyma kodoner för varje aminosyra, medan antikodoner är minst en och inte mer än tre.
Påminner om att gener är avsedda som enstaka sträckor av mycket långa polynukleotidsekvenser av DNA, är det klart att början och slutet av den enda genen nödvändigtvis måste finnas i minnet.
BIOSYNTES AV PROTEINER
I olika delar av DNA finns öppningen av dubbelkedjan och syntesen av de olika typerna av RNA.
Under laddningssteget binder RNAt till aminosyrorna (tidigare aktiverat av ATP och av det specifika enzymet). Det biosyntetiska "maskineriet" kan inte "korrigera" felaktigt laddade tRNA.
RNAr delas sedan upp i de två subenheterna och, genom att binda till de ribosomala proteinerna, ger upphov till sammansättningen av ribosomerna.
RNAm, som passerar genom cytoplasman, binder till ribosomerna och bildar polysomen.Varje ribosom, som flyter på budbäraren, är gradvis värd för RNAt som är komplementär till de relativa kodonerna, tar aminosyrorna och binder dem till polypeptidkedjan i bildning.
Det relativt stabila RNAt kommer in i cirkulationen igen. Ribosomerna används också igen och frigör den redan sammansatta polypeptiden.
Budbäraren, mindre stabil eftersom den är monokatenär, klyvs (av ribonukleaset) in i de konstituerande ribonukleotiderna.
Cykeln fortsätter således och syntetiserar efter varandra polypeptiderna på budbärar -RNA som tillhandahålls av transkriptionen.















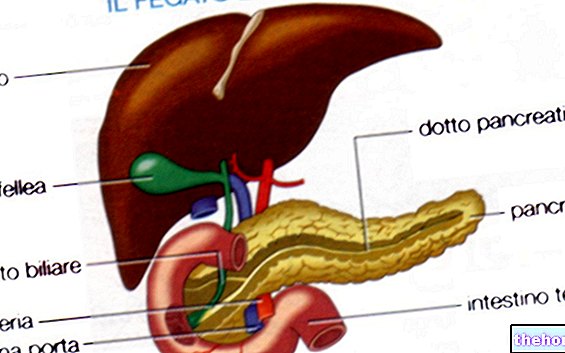



.jpg)

