För att kunna tala på ett begripligt sätt om "hemoglobin (Hb), det är användbart att ta hand om det första myoglobin (Mb) som är mycket lik hemoglobin men är mycket enklare.Mellan hemoglobin och myoglobin finns det nära släktskap: båda är konjugerade proteiner och deras protetiska grupp (icke-proteindel) är gruppen hem.
Myoglobin är ett globulärt protein som består av en enda kedja med cirka hundra femtio aminosyror (det beror på organismen) och dess molekylvikt är cirka 18 Kd.
Som nämnts är den utrustad med en hemgrupp som införs i en hydrofob (eller lipofil) del av proteinet, bestående av veck som kan hänföras till a-helixstrukturerna hos fibrösa proteiner.
Myoglobin består huvudsakligen av segment av α-helixer, närvarande i antal åtta och består nästan uteslutande av opolära rester (leucin, valin, metionin och fenylalanin) medan polära rester är praktiskt taget frånvarande (asparaginsyra, glutaminsyra, lysin och arginin); de enda polära resterna är två histidiner, som spelar en grundläggande roll i bindningen av syre till hemgruppen.
Hemgruppen är en kromoforgrupp (absorberas i det synliga) och är den funktionella gruppen av myoglobin.
Se även: glykat hemoglobin - hemoglobin i urinen
Lite kemi

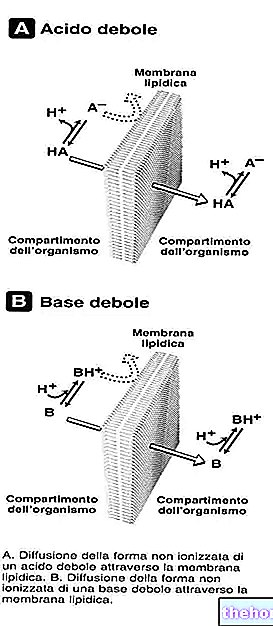
Bindningen mellan protoporfyrin och järn är en typisk bindning av koordinationsföreningar som är kemiska föreningar där en central atom (eller jon) bildar bindningar med andra kemiska arter i ett antal som är större än dess oxidationsnummer (elektrisk laddning). När det gäller hem är dessa bindningar reversibla och svaga.
Järnets koordinationsnummer (antal koordinationsbindningar) är sex: det kan finnas sex molekyler runt järnet som delar bindningselektronerna.
För att bilda en koordinationsförening krävs två orbitaler med rätt orientering: en kan "skaffa" elektroner och den andra kan donera dem.
I hem bildar järn fyra plana bindningar med de fyra kväveatomerna i mitten av proto-porfyrinringen och en femte bindning med ett proximalt histidinkväve; järn har den sjätte fria koordinationsbindningen och det kan binda till syre.
När järn är i form av en fri jon, dess typ orbitaler d de har alla samma energi; i myoglobin är järnjonen bunden till protoporfyrin och histidin: dessa arter stör magnetiskt orbitalerna d lite järn; störningen kommer att vara annorlunda för de olika orbitalerna d beroende på deras rumsliga orientering och den hos de störande arterna. Eftersom orbitalernas totala energi måste vara konstant, orsakar störningen en energisk separation mellan de olika orbitalerna: energin som förvärvas av vissa orbitaler motsvarar den energi som förloras av de andra.
Om avståndet mellan orbitalerna inte är särskilt stort, är ett elektroniskt arrangemang med hög snurrning att föredra: bindningselektronerna försöker ordna sig i parallella snurr i så många delnivåer som möjligt (maximal multiplicitet); om å andra sidan störningen är mycket stark och det finns en stor separation mellan orbitalerna kan det vara mer bekvämt att para ihop bindningselektronerna i orbitalerna med lägre energi (låg spinn).
När järn binder till syre antar molekylen ett lågt centrifugeringsarrangemang medan när järn har den sjätte koordinationsbindningen fri, har molekylen ett högt centrifugeringsarrangemang.
Tack vare denna snurrskillnad kan vi genom en spektralanalys av myoglobin förstå om syre (MbO2) är bunden till det eller inte (Mb).
Myoglobin är ett typiskt muskelprotein (men det finns inte bara i muskler).
Myoglobin extraheras från spermhvalen i vilken det finns i stora mängder och renas sedan.
Valar har en andning som hos människor: med lungor måste de absorbera luft genom andningsprocessen; spermhvalen måste föra in så mycket syre som möjligt till musklerna som kan ackumulera syre genom att binda det till myoglobinet som finns i dem; syret frigörs sedan långsamt när valen är nedsänkt eftersom dess metabolism kräver syre: desto större mängd syre som spermhvalen kan absorbera och desto mer syre är tillgängligt under dyket.
Myoglybin binder syre på ett reversibelt sätt och finns i perifera vävnader i en större andel desto mer vävnad är van vid att arbeta med syretillförsel som är avlägsna i tid.
<--- Myoglobin är ett protein som finns i musklerna, vars funktion är just en syre "reservoar".

Det som gör köttet mer eller mindre rött är halten av hemoproteiner (det är hemmen som gör köttet rött).
Hemoglobin har många strukturella likheter med myoglobin och kan binda molekylärt syre på ett reversibelt sätt; men medan myoglobin är begränsat till muskler och perifera vävnader i allmänhet, finns hemoglobin i erytrocyter eller röda blodkroppar (de är pseudoceller, det vill säga de är inte riktiga celler) som utgör 40% av blodet.
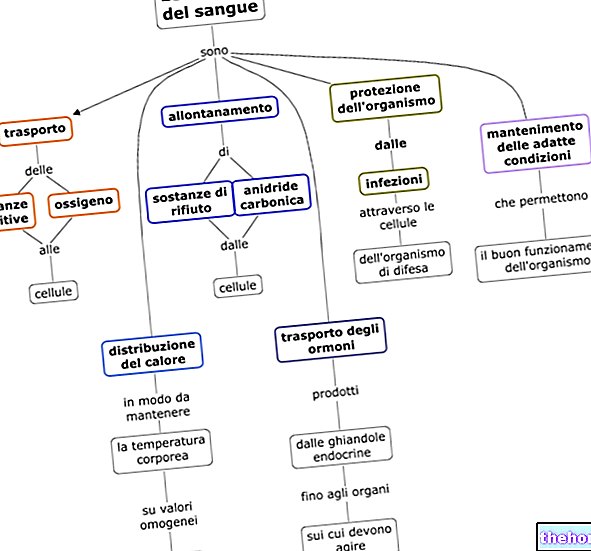
I motsats till myoglobin är hemoglobins arbete att ta syre i lungorna, släppa ut det i cellerna där det behövs, ta koldioxid och släppa ut det i lungorna där cykeln börjar igen.
L "hemoglobin det är en tetrameter, det vill säga den består av fyra polypeptidkedjor var och en med en hemgrupp och identiska två och två (i en människa finns det två alfakedjor och två betakedjor).
Hemoglobins huvudfunktion är transport av syre; en annan funktion av blodet i vilket hemoglobin är involverat är transport av ämnen till vävnaderna.
I vägen från lungorna (rik på syre) till vävnaderna, transporterar hemoglobin syre (samtidigt når de andra ämnena vävnaderna) medan det i den omvända vägen bär det avfall som samlas in av vävnaderna, särskilt kolet dioxid producerad i ämnesomsättningen.
I utvecklingen av en människa finns det gener som bara uttrycks under en viss tidsperiod; av denna anledning finns det olika hemoglobiner: fetala, embryonala, hos den vuxna mannen.
Kedjorna som utgör dessa olika hemoglobiner har olika strukturer men med vissa likheter är den funktion de utför mer eller mindre densamma.
En förklaring till förekomsten av flera olika kedjor är följande: under den evolutionära processen av organismer har till och med hemoglobin utvecklats som specialiserat sig på transport av syre från områden som är rika på det till områden som är bristfälliga. I början av evolutionskedjan l "hemoglobin transporterade syre i små organismer; under evolutionens gång nådde organismerna större dimensioner, därför modifierades hemoglobinet för att kunna transportera syre till områden längre bort från den punkt där det var rikt på det; till gör detta har de under den evolutionära processen kodats för nya strukturer i kedjorna som utgör hemoglobin.
Myoglobin binder syre även vid blygsamma tryck; i de perifera vävnaderna finns ett tryck (PO2) på cirka 30 mmHg: myoglobin vid detta tryck släpper inte ut syre, så det skulle vara ineffektivt som syrehållare. Hemoglobinet å andra sidan , det har ett mer elastiskt beteende: det binder syre till höga tryck och släpper ut det när trycket minskar.
När ett protein är funktionellt aktivt kan det ändra sin form lite; till exempel har syresatt myoglobin en annan form än icke-syresatt myoglobin och denna mutation påverkar inte sina grannar.
Situationen är annorlunda när det gäller associerade proteiner som hemoglobin: när en kedja syresätts induceras den att ändra form, men denna modifiering är tredimensionell så de andra kedjorna i tetrametern påverkas också. Det faktum att kedjorna är associerade med varandra., föreslår att modifieringen av en påverkar de andra grannarna även om det är i en annan utsträckning; när en kedja syresätter sig antar tetrameterns andra kedjor en "mindre fientlig inställning" till syre: svårigheten med vilken en kedja den syrehalten minskar när kedjorna nära den syrerar i tur och ordning. Detsamma gäller deoxygenering.
Deoxyhemoglobins kvartära struktur kallas T (spänd) form medan oxhemoglobin kallas R (frigjord) form; i det spända tillståndet finns det en rad ganska starka elektrostatiska interaktioner mellan sura aminosyror och basiska aminosyror som leder till en styv struktur av deoxyhemoglobin (det är därför den "spända formen"), medan när syre är länkat, är enheten av dessa interaktioner minskar (därav den "frigjorda formen"). Vidare, i frånvaro av syre, stabiliseras histidins laddning (se struktur) av den motsatta laddningen av asparaginsyran medan det i närvaro av syre finns en tendens hos proteinet att förlora en proton; allt detta innebär att syresatt hemoglobin är en starkare syra än deoxygenerat hemoglobin: bohr -effekt.
Beroende på pH binder hemgruppen mer eller mindre lätt till syre: i en sur miljö frigör hemoglobin lättare syre (den spända formen är stabil) medan bindningen till syre i en grundläggande miljö är svårare.

Varje hemoglobin släpper ut 0,7 protoner per mol syre (O2) som kommer in.
Bohr -effekten gör att hemoglobin kan förbättra förmågan att transportera syre.
Hemoglobinet som rör sig från lungorna till vävnaderna måste balansera sig själv som en funktion av tryck, pH och temperatur.
Låt oss se effekten av temperaturen.
Temperaturen i lungalveolerna är cirka 1-1,5 ° C lägre än den yttre temperaturen medan temperaturen i musklerna är cirka 36,5-37 ° C; när temperaturen ökar sjunker mättnadsfaktorn (vid samma tryck): detta händer eftersom kinetisk energi ökar och dissociation gynnas.

Det finns andra faktorer som kan påverka hemoglobins förmåga att binda till syre, varav en är koncentrationen av 2,3 bisfosfoglycerat.
2,3 bisfosfoglycerat är en metabolisk närvarande i erytrocyter i en koncentration av 4-5 mM (i ingen annan del av organismen finns det i en så hög koncentration).
Vid fysiologiskt pH deprotoneras 2,3 bisfosfoglycerat och har fem negativa laddningar på sig; den är fastklämd mellan de två betakedjorna av hemoglobin eftersom dessa kedjor har en hög koncentration av positiva laddningar. De elektrostatiska interaktionerna mellan betakedjorna och 2,3 -bisfosfoglyceratet ger systemet en viss styvhet: en spänd struktur erhålls som har liten affinitet för syre; under syresättning utvisas sedan 2,3 -bisfosfoglyceratet.
I erytrocyter är c "en speciell apparat som omvandlar 1,3 bisfosfoglycerat (producerat genom metabolism) till 2,3 bisfosfoglycerat så att det når en koncentration på 4-5 mM och därför kan hemoglobinet byta ut" syret i vävnaderna.
Hemoglobinet som anländer till en vävnad är i frigjort tillstånd (bundet till syre), men i vävnadens närhet karboxyleras det och passerar till det spända tillståndet: proteinet i detta tillstånd har mindre tendens att binda med syre, med respekt till frisläppt tillstånd, därför avger hemoglobin syre till vävnaden; Vidare sker det genom reaktion mellan vatten och koldioxid produktion av H + joner, därför ytterligare syre på grund av bohr -effekten.
Koldioxid diffunderar in i erytrocyten som passerar genom plasmamembranet; eftersom erytrocyter utgör cirka 40% av blodet, bör vi förvänta oss att endast 40% av koldioxiden som diffunderar från vävnaderna kommer in i dem, faktiskt 90% av koldioxiden kommer in i erytrocyterna eftersom de innehåller ett enzym som omvandlar koldioxid i kolsyra resulterar det i att den stationära koncentrationen av koldioxid i erytrocyterna är låg och därför är inmatningshastigheten hög.

Ett annat fenomen som uppstår när en erytrocyt når en vävnad är följande: genom gradient lämnar "HCO3- (derivat av koldioxid)" erytrocyt och för att balansera utmatningen av en negativ laddning har vi "kloridposten" som bestämmer en ökning av det osmotiska trycket: för att balansera denna variation finns det också inträde av vatten som orsakar svullnad av erytrocyt (HAMBURGER -effekt) Det motsatta fenomenet uppstår när en erytrocyt når lungalveolerna: en deflation av erytrocyterna (HALDANE -effekt) Därför är de venösa erytrocyterna (riktade till lungorna) rundare än de arteriella.















.jpg)

