ASSOCIATION (ELLER "CONCATENATION" ELLER "LINKAGE")
Hittills har vi behandlat Mendelian dihybrid (eller polyhybrid) korsningen med antagandet att de olika allelparen faktiskt finns på olika homologa kromosompar. Men antalet kromosomala par, även om det skiljer sig från art till art, varierar inom snäva gränser (få arter når runt hundra kromosomer), medan antalet gener kan räknas i tiotusentals.
Att karaktärerna som Mendel valde för sina experiment segregerade alla oberoende (utan att därför förväxla beräkningarna i fördelningen av fenotyper i F2 i polyhybridkorset) var en lycklig chans. Om två par alleler hittades på intilliggande loci, skulle lagen som skulle följa skulle kallas föreningslagen.
Att veta att väldigt många tecken har sin plats på en enda kromosom och att de är de kromosomala paren som separerar oberoende i meios, det kan mycket väl förstås hur ofta det händer att två par tecken, om de var associerade på en kromosom i föräldern organism, förbli associerad. lika mycket i könscellen och därför i den organism som den kommer att föra sitt eget genetiska material till.
Således ser vi att "förening representerar ett" undantag, långt ifrån sällsynt, från den självständighet som anges i Mendels tredje lag.
BYT ELLER "ÖVERKRIDNING" OCH KOMBINERING
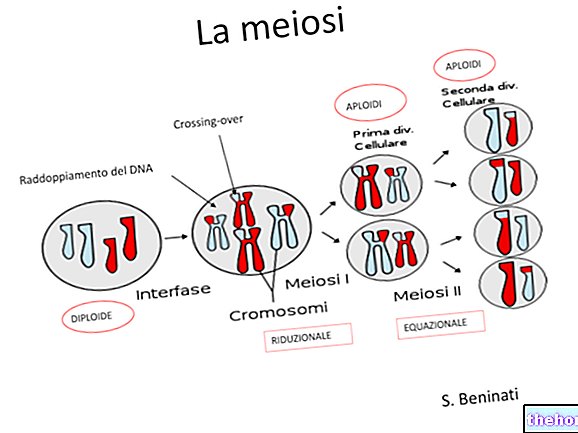
På tal om meios har vi indikerat att det finns två olika stunder för blandning av det genetiska materialet: en är den för segregeringen av kromosomer i könsceller, och det är den som observerats av Mendel.
Det andra ögonblicket, som faktiskt föregår, är det där de fyra kromatiderna i varje par homologa kromosomer ömsesidigt utbyter identiska egenskaper. Efter denna utbyte kommer två faktorer som var associerade på samma kromosom istället att vara oberoende i könscellerna. Sannolikheten att ett utbyte äger rum är proportionellt, med en första approximation, till kromosomens längd, och i längre kromosomer kan det bli ännu mer än, ett utbyte.
Fenomenet kan detekteras cytologiskt genom att observera ett tillräckligt antal meioser under mikroskopet.
Åter matchningsfrekvensen är den hastighet med vilken två tecken som var associerade i föräldragenerationen rekombinerar annorlunda i F2.
Om de två loci är absolut sammanhängande är sannolikheten att en chiasm kommer att skilja dem praktiskt taget noll. Rekombinationshastigheten kommer att vara: n ° rekombinanter. Om två loci finns på två olika kromosomer, kommer rekombinationshastigheten att vara 0,5 (lika sannolikhet, för två tecken som var sammanfogade i P -generationen, för att slumpmässigt hitta ihop i F2). Rekombinationshastigheten kan därför variera mellan 0,0 och 0,5. För små avstånd på kromosomen är avståndet och rekombinationshastigheten direkt proportionell. För längre sträckor finns det möjlighet att två utbyten sker mellan två platser. Det kommer nu att framgå som klart att två faktorer som separeras av två börser återigen är associerade. det är klart, vid denna tidpunkt, att proportionaliteten mellan avståndet till loci och sannolikheten för rekombination förloras.
Loci som finns associerade på samma kromosom utgör 'föreningsgrupper'. Mycket avlägsna platser kan ha en sådan sannolikhet för separation genom utbyte att de beter sig som oberoende, men var och en av dem kommer att associeras, med en lägre rekombinationshastighet, till mellanliggande loci.
När rekombinationshastigheterna mellan många par av gener inom en föreningsgrupp är kända kan konstruktionen av "genetiska kartor" börja. Med tanke på att avståndet mellan två gener (a och b) uttrycks av rekombinationshastigheten och att avståndet från a från en tredje gen c kan vara antingen summan eller skillnaden med avståndet från b, är det möjligt att rekonstruera en karta över de ömsesidiga avstånden, som kommer att vara den genetiska kartan inom den gruppen av föreningar, det vill säga den kromosomen.
Vi måste nu överväga några begrepp som begränsar den fenotypiska manifestationen av genotypiska karaktärer.
Först och främst kommer vi att prata om begreppen penetration och uttrycksfullhet, och sedan kommer vi att ägna särskild uppmärksamhet åt fenomenen reglering av genverkan.
PENETRANS
Genets penetration representerar dess förmåga att manifestera sig i fenotypen. Penetrance mäts statistiskt genom att räkna frekvensen av fenotyperna som visar den karaktären av 100 genotyper som innehåller den. En egenskap med 0,7 penetration är en egenskap som förekommer fenotypiskt i 70% av dess genotypiska frekvens.
EXPRESSIVITET
Expressivitet är en kvantitativ bedömning av graden av fenotypisk manifestation.
REGLERING OM ALLMÄNNA ÅTGÄRDER
Celler producerar alla sina enzymer och proteiner vid samma hastighet och samtidigt. Escherichia coli-celler kan till exempel förses med energi och kolatomer från laktosdisackariden eftersom de kan bryta ner dem till glukos och galaktos tack vare beta-galaktosidasenzymet. I en normal E. coli som kan ha laktos, det finns ungefär 3 000 molekyler av beta-galaktosidas, lika med 3% av proteinerna i den cellen; i frånvaro av laktos kommer det bara att finnas en molekyl av beta-galaktosidas per bakteriecell. Galaktosidas kommer att syntetiseras från nya mRNA -molekyler när det kan användas. Mutanta stammar av E. coli som är rika på enzymet är kända även när laktos saknas: dessa mutanter missgynnas jämfört med normala celler eftersom de tvingas till onödig förbrukning av energi och material för att producera enzymet som kommer att förbli utan substrat. Ämnen som orsakar en ökning av mängden enzym, som är fallet med laktos, kommer att kallas inducerare, medan enzymer sägs vara inducerbara. Andra ämnen inducerar, även dessa på ett specifikt sätt, produktionen av vissa enzymer. Även i E. coli, till exempel, som kan bygga alla dess aminosyror, med kol och ammonium (NH3), blockerar närvaron i odlingsmediet av en viss aminosyra (till exempel histidin) produktionen av alla enzymer associerade med aminosyrans biosyntes: det kommer att sägas om dessa enzymer att de är repressiva. I bakterieceller rivs mRNA -molekylerna strax efter bildandet, och det är därför som kontroll av produktionen av mRNA innebär att man samtidigt kontrollerar den enzymatiska syntesen tid.
OPERONEN
För att förklara hur bakteriecellen kan styra sin egen produktion av enzymer formulerade Jacob och Monod hypotesen om operon; operonen bildas av flera gener som är funktionellt besläktade och inriktade utan diskontinuitet längs en sträcka av DNA. Operonen består av tre olika typer av gener: promotorn, där bildandet av mRNA börjar; operatören, där kontroll utövas, en eller flera strukturgener, som kodar för enzymer eller för andra proteiner. I betagalaktosidasystemet inkluderar operon, förutom det för betagalaktosidas, även två andra gener som strukturellt kodar för andra enzymer som är involverade i metabolismen av laktos. Dessa gener ligger intill varandra och transkriberas efter varandra längs samma DNA -spiral till en enda mRNA -molekyl. De sålunda producerade mRNA -molekylerna är aktiva under mycket kort tid, varefter de förstörs av specifika enzymer.
Operonens aktivitet styrs i sin tur av en annan gen, regulatorn, som också kan vara avlägsen operonen: denna regulator kodar för ett protein, kallat en repressor, som verkar binda till DNA vid operatörsgenen. mellan promotorn och de strukturella generna blockerar faktiskt produktionen av mRNA.
Repressorn styrs i sin tur och kontrollen utförs med hjälp av en "signal" substans. När det gäller inducerbara enzymer är detta ämne "induceraren. Induceraren" binder till repressormolekylen som ändrar dess form så att den inte längre kan anpassa sig till DNA: i detta fall, eftersom det inte finns någon repressor mellan promotorn och strukturgener kan repressorn bilda mRNA -molekylerna och från dessa proteinmolekylerna. När förbrukaren av inducerare har tömts igen kommer regulatorn att återfå kontrollen, vilket kommer att stoppa produktionen av nytt mRNA, därför av nya proteiner.I beta-galaktosidasystemet är induceraren laktos eller ett ämne som liknar detta. Derivat: de kommer att ansluta sig till repressorn och inaktivera den för att möjliggöra biosyntes av enzymer. När det gäller repressiva enzymer fungerar ämnet som fungerar som en "signal" som en corepressor: repressorn är endast aktiv om den kombineras med corepressorn. I histidinsystemet, som involverar ett dussin olika enzymer, är det denna aminosyra, i kombination med dess tRNA, corepressor. Histidin.
ALLOSTERISKA INTERAKTIONER
Allosteriska interaktioner, som involverar inaktivering av ett enzym genom att ändra dess form, ger ett annat sätt att reglera cellens metaboliska aktivitet. Allosteriska interaktioner möjliggör mer exakt kontroll än operonets induktor -repressorsystem, men uppnår inte det användbara resultatet att utesluta biosyntesen för ett givet ämne från det första steget - produktionen av ett mRNA.
KONTROLLSYSTEM I EUCARIUS
Det finns några fakta som leder till att man tror att ett regleringssystem som liknar operon fungerar och är framträdande bland växter och djur.Kromosomerna hos dessa organismer skiljer sig kraftigt från E. coli och andra prokaryoter. Kontrollen av generna i dessa cellerna är mycket olika. Mitosmekanismen är sådan att varje cell i en viss växt eller ett djur besitter all information
genetik som finns i det befruktade ägget. Därför kommer de flesta generna i någon specialiserad cell att förbli ineffektiva under cellens hela livslängd. DNA: t i dessa celler är alltid associerat med proteiner. Så det är möjligt att genförträngning i eukaryoter kräver just detta samband mellan DNA och proteiner.