Utan genetisk variation bör alla levande saker (genom arv) vara desamma som de första. För att ha ojämlika varelser skulle de enda förklaringarna vara de som rör enskilda skapelser. Men vi vet att DNA -strukturen, som är grunden för överföring av ärftliga karaktärer, har en relativ och inte en absolut stabilitet. Medan stabiliteten garanterar bevarandet av den initiala informationen, bestämmer instabiliteten modifieringarna, eller bättre (för att använda den specifika termen) mutationerna.
Mutationer kan delas in i tre stora grupper:
- genmutationer;
- kromosomala mutationer;
- genomiska mutationer.
Vid denna tidpunkt är det bra att kort lägga till två begrepp: det ena är "reparation" och det andra är "gunstig" eller "ogynnsam" mutation.
Reparationen, en engelsk term som betyder reparation, avser förekomsten i cellerna av specifika enzymatiska processer avsedda att verifiera bevarandet av information, eliminera eller korrigera DNA -sträckor som inte överensstämmer med originalet.
Begreppet "gynnsam" eller "ogynnsam" mutation uttrycker jämförelsen mellan effektiviteten, för organismen och för arten, av den ursprungliga genen ("vild", dvs "vild", primitiv) och av mutantgenen.
Det bör noteras att fördelar och nackdelar alltid avser vissa miljöförhållanden; en ogynnsam mutation i en viss miljö kan bli gynnsam under olika förhållanden.
Konsekvenserna av denna princip är enorma inom populationsgenetik, vilket redan kan demonstreras av de olika överlevnadseffektiviteterna av hemoglobinvarianter i normala eller malariala miljöer. Men detta ämne bör skjutas upp till mer detaljerade artiklar om allmän, mänsklig eller medicinsk genetik.
En annan skillnad krävs (särskilt för genmutationer) mellan somatiska mutationer och könsmutationer. Alla celler i vår flercelliga organism härrör som bekant från en enda zygot, men väldigt tidigt skiljer de sig till en somatisk linje, från vilken hela kroppen utvecklas, och en könslinje avsedd att bilda könskörtlarna och, för terminal meios, gameterna Det är ganska uppenbart att, med undantag för interaktioner mellan de två cellpopulationerna, kommer en mutation av den somatiska linjen att manifestera sig i den enda organismen men kommer inte att påverka ättlingarna, medan en mutation i könscellerna endast kommer att inträffa hos ättlingarna.
När det gäller de olika typerna av mutation och de relativa konsekvenserna är det värt att återigen nämna en klassificering av orsakerna till mutation. Dessa orsaker kallas mutagener, som används för att skilja huvudsakligen till fysiska och kemiska mutagener. Olika modifieringar av den fysiska miljön kan leda till mutationer, men de huvudsakliga fysiska mutagenerna är strålning. Av denna anledning är radioaktiva ämnen farliga, och det är framför allt de tyngre radioaktiva partiklarna som tenderar att orsaka mutationer genom avhopp, med de allvarligaste konsekvenserna.
Kemiska mutagener kan verka både genom att ändra den ordnade strukturen för nukleinsyror och genom att introducera ämnen som liknar normala kvävehaltiga baser i cellerna, som kan konkurrera med de senare i syntesen av nukleinsyror, vilket orsakar mutationer genom substitution.
GENE MUTATIONER
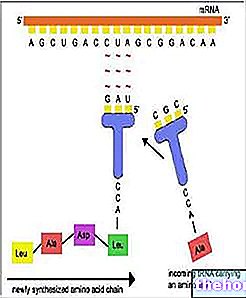
Genmutationer påverkar en eller några gener, det vill säga en begränsad sträcka av DNA. Eftersom information lagras i en sekvens av nukleotidpar, består den minsta mutationsenheten (ett muton) av ett enda komplementärt baspar. Utan att gå in på detaljer om de olika mutationsmekanismerna på gennivå kan vi begränsa oss till att nämna två: basersubstitution och omval (eller infogning). Vid basersubstitutionsmutationer ersätts en eller flera DNA -nukleotider med andra. Om felet inte korrigeras i tid kommer en sekvens att följa vid transkriptionstillfället också förändras i RNA. Om ändringen av triplet är inte begränsat till en synonym (se den genetiska koden), det kommer att resultera i substitution av en eller flera aminosyror i polypeptidsekvensen. Substitutionen av en aminosyra kan vara mer eller mindre kritisk för bestämning av proteinets struktur och dess funktion.
Vid omval eller insertionsmutationer avlägsnas eller adderas en eller flera nukleotider till DNA -sekvensen. Dessa mutationer är i allmänhet mycket allvarliga eftersom (om de inte är hela trillingar som adderar eller subtraherar enstaka aminosyror) ändras alla trillingar som följer i läsordningen.
Genmutationer är de vanligaste och är ursprunget till de flesta variabiliteten av ärftliga egenskaper mellan individer.
KROMOSOMISKA MUTATIONER
Dessa är mutationer som påverkar relativt långa fragment av en kromosom. Det är vanligt att klassificera dem huvudsakligen i:
- kromosomala mutationer för omval;
- kromosomala mutationer genom dubbelarbete;
- kromosomala mutationer genom translokation.
Mutationer på grund av avhopp uppstår på grund av brott och förlust av ett mer eller mindre långt fragment av kromosom. Särskilt vid meios är denna typ av mutation ofta dödlig, på grund av den totala förlusten av ett visst antal gener, mer eller mindre oumbärlig.
Vid dubbelmutationer, efter en paus, tenderar kromatidstubbarna att smälta samman.
I det efterföljande avlägsnandet av centromererna bryts kromosomen, som har blivit dicentrisk, i ofta ojämlika delar: som vi kan se är resultatet omval på ena sidan och dubblering å andra sidan.
En kromosombrytning kan följas av en vändning. Det totala genetiska materialet är oförändrat, men sekvensen av generna på kromosomen ändras.
Fallet med en translokation är liknande, men det gäller bindningen av ett fragment av en kromosom till en icke-homolog kromosom. En kromosom amputeras och den andra förlängs; cellens totala genetiska information är fortfarande oförändrad, men positionseffekten är ännu mer markant.Det är lätt att representera en positionseffekt genom att hänvisa till begreppet reglering av genverkan: genom att ändra position på kromosomer, kan en gen enkelt lämna en operon och sättas in i en annan, vilket resulterar i en "förändrad aktivering eller repression.
Det sägs emellertid att en translokation är balanserad (eller balanserad) när det finns ömsesidighet av translokation mellan två par kromosomer, vilket håller summan av geninformationen oförändrad. Den korsformade figuren i meiotisk diakinesis motsvarar i allmänhet den balanserade translokationen.
GENOMISKA MUTATIONER
Med tanke på att genomet är det individuella genetiska arvet, ordnat i kromosomer, kan det specificeras att vi talar om genomiska mutationer när kromosomerna har en annan fördelning än artens norm.
Genomiska mutationer kan huvudsakligen klassificeras i mutationer för polyploidi, haploidi och aneuploidi.
Polyploidimutationer uppstår när reduktion inte leder till delning; de förekommer lättare i växter, där de verkligen används för att förbättra produktionen.
Om celldelningen misslyckas vid meios kan diploida gameter förekomma; om en sådan könscykel lyckas smälta samman med en normal könscykel, blir zygoten från denna befruktning triplo. En sådan zygot kan ibland lyckas härleda en hel organism, eftersom reduktion och mitos inte kräver ett jämnt antal kromosomer. Vid tiden för meios är det emellertid omöjligt att regelbundet para homologa kromosomer.
Genomiska mutationer av haploidi kan inträffa när en gamete i en normalt diploid art aktiveras av en annan könscykel utan kärnmaterial, eller till och med i frånvaro av befruktning: en haploid individ kommer att uppstå.
Medan de tidigare typerna av genomiska mutationer alltid gäller addition eller subtraktion av heltal n av kromosomer, gäller mutationer för aneuploidi överskott eller defekt hos enstaka kromosomer (kromosomavvikelser).
Den euploida kromosomala sammansättningen av en art definieras som dess normala karyotyp eller idiotyp.






-perch-e-quando-si-misura.jpg)









.jpg)